
Ectothermic animals are strongly influenced by temperature. It is speculated that species face a trade-off between increased performance at a specific temperature versus being able to function over a broader range of temperatures but at a lower peak level. The balance of this trade-off – if it exists – may influence the environments that fish and other ectotherms inhabit and how they respond to climate change. A new paper by Julie Nati, Jan Lindstrom, Lewis Halsey and Shaun Killen shows that, across fish species, peak performance for aerobic scope is not linked to performance across a range of temperatures, suggesting that links between peak and thermal breadth for the capacity for aerobic metabolism may not affect responses to climate warming. Read more in on access paper here!:
http://rsbl.royalsocietypublishing.org/content/12/9/20160191
Nati, JH, Lindstrom, J, Halsey, LG, & Killen, SS. 2016. Is there a trade-off between peak performance and performance breadth across temperatures for aerobic scope in teleost fishes? Biology Letters. DOI: 10.1098/rsbl.2016.0191
Viewing entries tagged
metabolism

Many animal species live in groups to derive benefits for foraging and predator avoidance. Another potential benefit is a reduction in routine energy expenditure for each individual within the group as they share duties for anti-predator vigilance. There may also be a ‘calming effect’, whereby individuals in group experience decreased stress and energy expenditure compared to isolated individuals. In a new study on the cover of this month’s Journal of Experimental Biology, Shaun Killen, Lauren Nadler, and colleagues at James Cooke University in Australia examined these issues by measuring metabolic and growth rates in individual damselfish (Chromis viridis) with and without visual and olfactory cues from groups of other fish of the same species. They found that individuals in shoals reduced their metabolic rate by 26% from their metabolic rate when alone. As increased extreme weather events may lead to forced social isolation in gregarious fishes, this could have repercussions for individual energy budgets. Photo credit: Eva McClure. Read more in the open access article here!:
http://jeb.biologists.org/content/219/18/2802
Nadler, L.E., Killen, S.S., McClure, E.C., Munday, P.L., McCormick, M.I. 2016. Shoaling reduces metabolic rate in a gregarious coral reef fish species. Journal of Experimental Biology. 219: 2802-2805. doi: 10.1242/jeb.139493

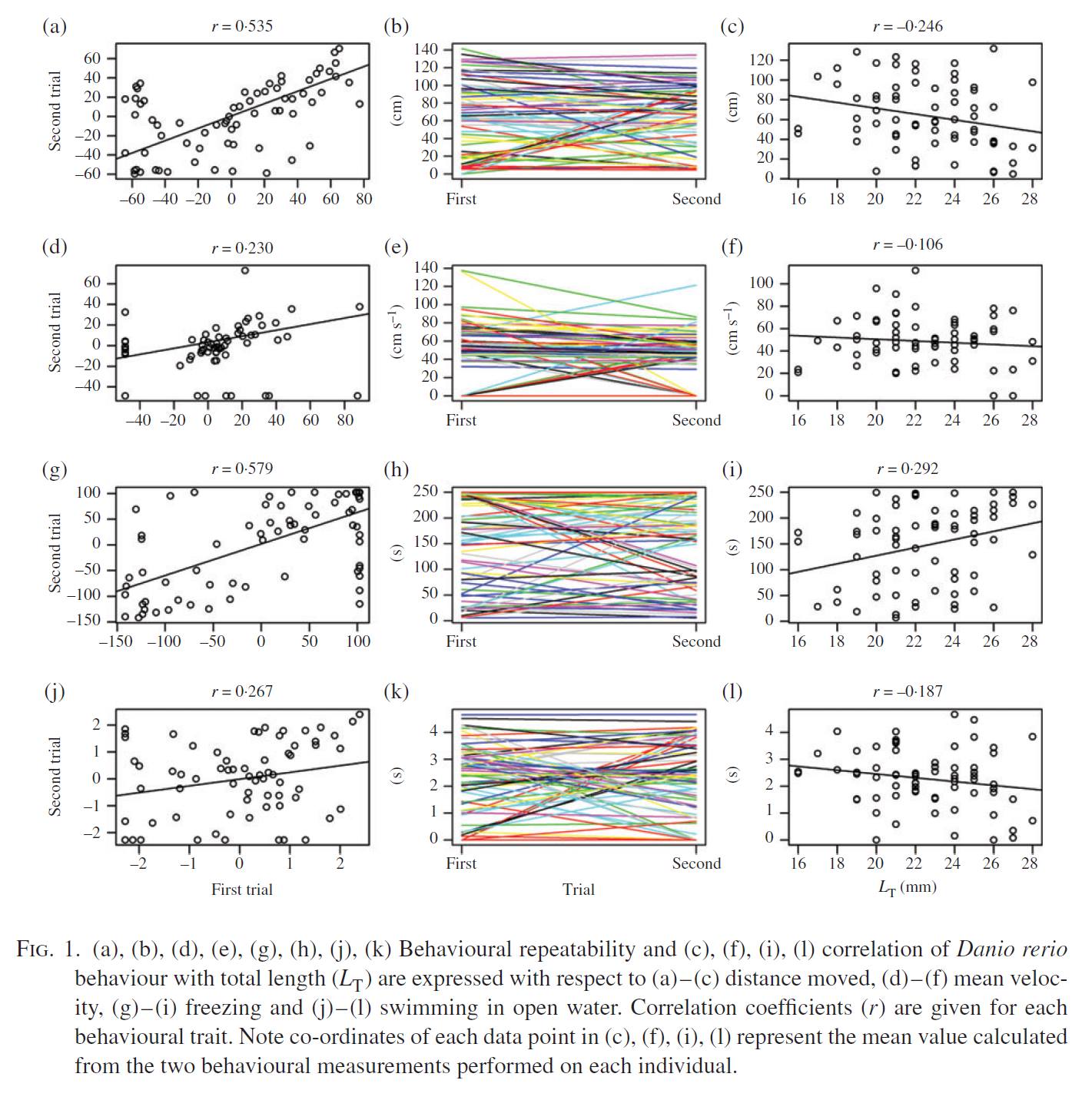
Individuals within animal species often show wide variation in behavioural traits, such as willingness to explore novel habitats or take risks around predators. This behavioural variation is often correlated with other traits including body size or metabolic rate. In a new study by Shaun Killen and colleagues in Germany, it was observed that for juvenile zebrafish, the tendency of individuals to take risks was explained by variation in body length but not necessarily body mass or baseline metabolic demand.
Read more in the open access article in the Journal of Fish Biology, here!:
http://onlinelibrary.wiley.com/doi/10.1111/jfb.13100/full
Polverino, G., Bierbach, D., Killen, S.S., Uusi-Heikkilä, S., Arlinghaus, R. 2016. Body length rather than routine metabolic rate and body condition correlates with risk-taking behaviour in juvenile zebrafish (Danio rerio). Journal of Fish Biology. doi:10.1111/jfb.13100

Sexual reproduction is often viewed as a cooperative interaction that passes on the genes of individuals while also propagating the species. In most species, however, there exists some form of sexual conflict with a disparity in the reproductive interests between females and males. For example, male reproductive success is typically limited by access to females. Males of many species try to overcome this using a number of behaviours, such as chasing and even attacking females in an attempt to gain a mating.
These types of behaviours are considered sexually harassing as males are attempting to coerce females into mating with them. Females can spend a lot of energy avoiding males in these situations and can even be injured. To reduce these costs, one possibility is that females may be able to change their own behaviour or physiology in ways that reduce the negative energetic consequences of harassment, or allows them to more easily escape male coercion.
In a recent paper, performed with the fantastic Darren Croft and Safi Darden at the University of Exeter, we studied this likelihood in a laboratory setting by housing female Trinidadian guppies (Poecilia reticulata) for several months with varying levels of male harassment that they would normally encounter in the wild. In nature male guppies spend a large portion of their time chasing and harassing females in an attempt to mate with them. Females can attempt to avoid this harassment by rapidly swimming away from the male during pursuits. After the five months of holding we then compared the swimming ability and energy spent while exercising in the females guppies using swim tunnel respirometry.
We had originally suspected that females exposed to more chasing may develop faster maximum swim speeds or a greater aerobic capacity. Basically, we thought that the constant chasing that they receive might induce a sort of training effect whereby the females would become better swimmers. After five months, however, females exposed to the varying levels of harassment showed no differences in either maximum swim speed or peak aerobic metabolism. But, what we DID see was a surprising increase in the efficiency of movement: females that had received high levels of harassment were able to swim much more efficiently, using less energy to swim at a given speed compared to those exposed to lower levels of harassment:

Figure 1. Changes in the oxygen required to swim a various speeds in female guppies. White circles represent females that had been exposed to high levels of male harassment for five months; black circles are females receiving low levels of harassment. From Killen, S.S., Croft, D.P., Salin, K., Darden, S.K. 2015. Male sexually coercive behaviour drives increased swimming efficiency in female guppies. Functional Ecology. doi: 10.1111/1365-2435.12527
This change in the efficiency of movement over time was unexpected and as far as we know has not been previously demonstrated in animals in response to an ecological pressure (here, sexual harassment). Luckily, my colleague Lewis Halsey turned us on to some of the relevant human exercise physiology literature. Amazingly, the changes in swimming efficiency displayed by our guppies bare a striking resemblance to the improvements made by human athletes in response to training. Compare the figure above to the figure below, which shows data for human runners:

Figure 2. Changes in the oxygen used for running various speeds in human athletes, before and after 6 weeks of endurance training. From Jones, A. & Carter, H. (2000) The effect of endurance training on parameters of aerobic fitness. Sports Medicine, 29, 373–386.
Even for humans it’s not exactly clear how this increase in efficiency with training is achieved. In our study, we found no effect of harassment level on the concentration of aerobic enzymes present within the muscles used for swimming. It is also possible that differences in mitochondrial efficiency play a role, but this is an area which will require further study. Another possibility is that there are changes to the mechanics of swimming that increase efficiency. This would be similar to changes in gait or posture while running or performing some other type of exercise. Indeed, we observed that females that experienced lower levels of harassment spent more time swimming with their pectoral fins extended, which previous studies have shown is an indicator of an inefficient swimming technique in guppies.
Overall, it seems that prolonged increases in high-intensity swimming in females, caused by male harassment, leads to changes in the physiology or swimming mechanics of individual fish, which reduce costs of swimming. Increased swimming efficiency or performance could also allow female guppies to escape male coercion more easily, giving them more control over matings. An exciting opportunity now exists to examine the extent to which this phenomenon occurs in the wild. This work is also an interesting example of how fundamental biological research can yield unexpected insight into the parallels between animals and humans: in this case, how human exercise physiology mirrors the plasticity displayed by animals in response to ecological pressures.